Imm02
Limfocyty 2.9
• Rozpoznanie antygenu przez powierzchniowe receptory
wiążące antygen.
• Przeciwciała monoklonalnc przeciw- CD3-TCR lub przeciw jednemu z epitopów CD2.
• Lektyny.
Lektyny są białkami, otrzymywanymi z roślin i bakterii, wiążącymi węglowodany. Niektóre z nich są zdolne do aktywacji limfocytów przez krzyżowe wiązanie z BC'R lub TC'R i są znane jako mitogeny (induktory proliferacji). Stymulacja limfocytów mitogenem in vitro jest uważana za dość dobrą pozorowaną stymulację limfocytów przez antygen. Komórki T i B są aktywowane różnymi mitogenami. Fitohcmaglutynina (PI IA) i konkanawalina A (Con-Aj stymulują ludzkie i mysie komórki T. Lipopolisacharyd (LPS) stymuluje mysie komórki B, podczas gdy mitogen szkarłatki (PWM) stymuluje zarówno ludzkie komórki T, jak i B (Ryc. 2. IR).
Użycie tych czynników in vitro wykazało, że aktywacja komórek T i B wywołuje produkcję cytokin i ich receptorów , które razem prowadzą wyselekcjonowane klony poprzez ich cykl życiowy (proliferacja) ku dojrzew aniu i produkcji komórek efek-torowych lub komórek pamięci (palrzRyc. 1.14 i 1.20). Tc ostatnie recyrkulują i ostatecznie umiejscawiają się w przestrzeniach T- lub B-zalcżnyeh tkanek limfoidalnych, gdzie pozostają gotow-c do odpowiedzi, jeżeli spotkają się znowu z tym samym antygenem.
Sygnał aktywacyjny jest przekazywany przez „wtórne przekaźniki"(„second messengers")
Współdziałanie spoczynkowego limfocyta z antygenem daje początek pewnej liczbie wczesnych zdarzeń biochemicznych, które powodują wytworzenie „wtórnych przekaźników” wewnątrz komórki B lub T. Przekaźniki są odpowiedzialne za zmiany na poziomic DNA komórki. Zarówno komórki T jak i B zużytkowują GTP-zależny komponent (lub białko G), aby wytworzyć reakcje sygnałowe , które oddziaływują przez stymulowanie przemiany fosfatydyloinozytolu. Reakcja ta wytwarza dwa wtórne przekaźniki, 1,4,5-trójfosforan inozytolu (IPV) i diacylo-gliccrol. IPi wywołuje uwalnianie Ca2' z wewnętrznych zasobów, a diacyloglicerol aktywuje kinazę białkową C. Razem z innymi kinazami, kinaza białkowa C fosforyluje pewną ilość powierzchniowych cząsteczek, prowadząc do aktywacji określonych genów. Wkrótce po połączeniu limfocytów T z antygenem. pojawia się pewna ilość powierzchniowych cząsteczek, takich jak gp39 i receptory dla cytokin, jak IL-2. Współdziałanie z tymi cząsteczkami powoduje proliferację i dojrzewanie limfocytów.
Różnicowanie komórek B prowadzi do tworzenia komórek plazmatycznych i komórek pamięci
W następstwie aktywacji komórek T i B przez mitogen lub antygen, obserwuje się wyraźne cechy różnicowania na poziomie ultrastrukturalnym (patrz Ryc. 2.8 i 2.19). Ostatecznie, dużo blastycznych komórek B dojrzewa do komórek tworzących przeciwciała (amibody-forming cclls, AFC). które przekształcają się dalej in vivo w końcowo zróżnicowane komórki plazmatycznc. Niektóre blasty B nie rozwijają szorstkich cndoplazmalycznych cystern siateczki. Komórki te znajdują się w ośrodkach rozmnażania i nazywają się komórkami centrum grudki lub centrobla-stami i centrocytami. Wydaje się więcej niż prawdopodobne, że są one są komórkami B pamięci, których istnienie jest pochodną zdolności układu immunologicznego do rozwoju trwalej odporności (Ryc. 2.20).
W mikroskopie świetlnym cytoplaznta komórek plazmatycznych jest zasadochłonna; jest to spowodowane dużą ilością RNA zużywanego do syntezy przeciwciał w szorstkiej siateczce śród-plazmatycznej (Ryc. 2.21). Na poziomie ultrastrukturalnym, szorstka siateczka śródplazmatyczna często jest widoczna w ułożeniu równoległym (Ryc. 2.22). Komórki plazmatyczne rzadko spotyka się w krążeniu, ponieważ stanowią one mniej niż 0,1% całkowitej populacji limfocytów. Normalnie są ograniczone do wtórnych narządów limfatycznych i tkanek. Przeciwciała produkowane przez jedną komórkę plazmatyczną mają tę samą swoistość i klasę Ig.. Immunoglobuliny można uwidocznić w cytoplazmie komórki plazmałycznej za pomocą reakcji ze swoistymi przeciwciałami znakowanymi fluorochromem (Ryc. 2.23). Komórki plazmatyczne mają krótki okres życia, trwający tylko kilka dni; giną w wyniku apoptozy (Ryc. 2.24).

Ryc. 2.18. Blastogeneza limfocytów wywołana mitogenem/antygenem. Widoczne ludzkie komórki T i komórki B, stymulowano mitogenem szkarłatki. Na lewo: Występuje zwiększona zasadochlonność w cytoplazmie i wzrost objętości komórki. Na prawo: Chromosomy zagęszczają się podczas podziału komórki i są wyraźnie widoczne podczas metafazy. Barwienie met. Giemsa, x2000.

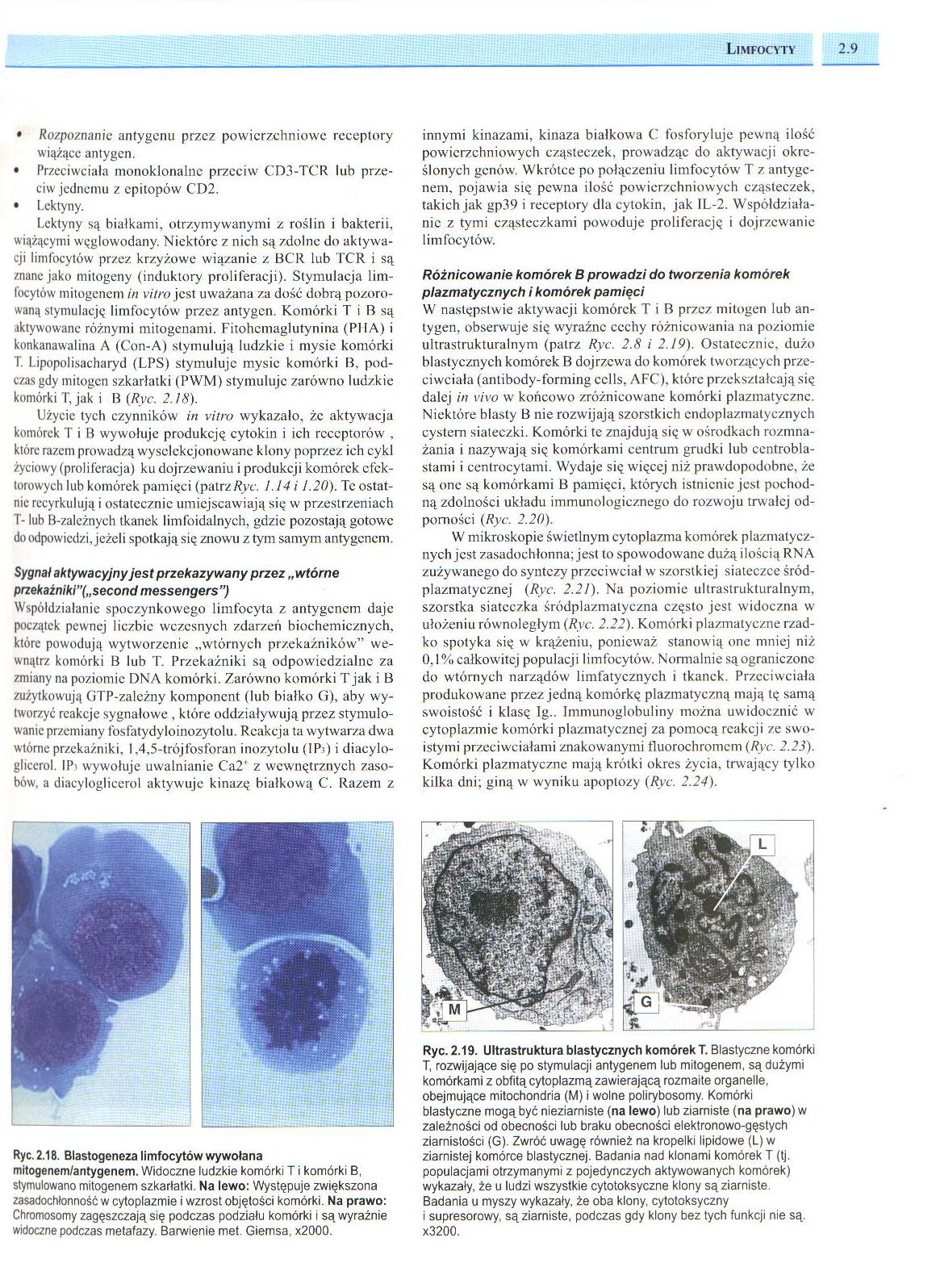
Ryc. 2.19. Ultrastruktura blastycznych komórek T. Blastyczne komórki T, rozwijające się po stymulacji antygenem lub mitogenem, są dużymi komórkami z obfitą cytoplazmą zawierającą rozmaite organelle, obejmujące mitochondria (M) i wolne polirybosomy. Komórki blastyczne mogą, być nieziarniste (na lewo) lub ziarniste (na prawo) w zależności od obecności lub braku obecności elektronowo-gęstych ziarnistości (G). Zwróć uwagę również na kropelki lipidowe (L) w ziarnistej komórce blastycznej. Badania nad klonami komórek T (tj. populacjami otrzymanymi z pojedynczych aktywowanych komórek) wykazały, że u ludzi wszystkie cytotoksyczne klony są ziarniste.
Badania u myszy wykazały, że oba klony, cytotoksyczny i supresorowy. są ziarniste, podczas gdy klony bez tych funkcji nie są. x3200.
Wyszukiwarka