48153 skanowanie0047 (22)

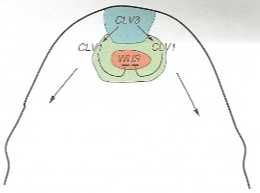
Rys. 15-15. f\xtowemhof<aoędunadare»yclB-presji genów CLAVATAś. CLAVATA< oraz WJSCHSL kontroiuiących wzajemnie s*ą s<lywność w pętfi sprzężenia znrołnegc i gwarantując^ w ten sposób odnawianie się konórek twó«czych na szczycie wierz-chdka oraz kierowanie ich pochodnych na ścieżkę rtórBCOasiTE
chołck kończy przedwcześnie swą działalność, różnicując na swej powierzchni wierzchołki przybyszowe, których rozwój jest ponownie wstrzymywany przez różnicowanie (rcitcracja programu rozwojowego). Wierzchołki kwiatowe wuschel kończą wzrost na inicjacji pręcika położonego terminalnie. Badania chimer peryklinalnych, w których tylko pierwsza warstwa tuniki (Li) miała komórki zdolne do ekspresji CLV3, pokazały, że to wystarczy do prawidłowego funkcjonowania wierzchołka.
15-3.1.2.3. Merystem generatywny
Najbardziej fascynującym, a jednocześnie jednym z najwcześniej opisanych systemów segmentacji w kierunku apikalno-bazalnym jest system ABC odpowiedzialny za określanie tożsamości organów kwiatowych. System len został odkryty w wyniku analizy fenotypów mutantów homeotycznych dwóch roślin modelowych: Arabidopsis iAniirrhi-num. Do określenia tożsamości czterech kolejnych segmentów kwiatowych: działek kielicha, płatków korony, pręcików i owocolistków konieczna jest aktywność trzech klas genów homeotycznych. Geny klasy A są aktywne na poziomie pierwszym i drugim, klasy B - na drugim i trzecim, a klasy C-na trzecim i czwartym. Ze względu na nakładanie się na siebie domen ekspresji genów A i B oraz B i C jest możliwe nadanie tożsamości aż czterem odrębnym typom organów kwiatowych. Obecność produktów ekspresji genów klasy A na poziomie pierwszym określa tożsamość działek kielicha. Na poziomie drugim rozpoczyna się ekspresja genów klasy B, połączona aktywność produktów genów klas A i B nadaje zawiązkom tożsamość płatków korony. Na poziomie trzecim nudal trwa ekspresja genów klasy B, ale zostają wyłączone geny klasy A, co oznacza automatyczne włączenie genów klasy C (A i C hamują się wzajemnie). Produkty genów B i C w tym samym obszarze określają tożsamość pręcikowia. Wyłączenie genów B w czwartym segmencie, a więc obecność tylko aktywności C, oznacza wytwarzanie owocolistków i jednocześnie zakończenie organogenczy (rys. 15-16). Geny klasy C są również odpowiedzialne za zdeterminowany wzrost pędu gcncraiywncgo. W domenie 0 zostaje też wyłączony gen WUS.
Wzór ekspresji genów dla kolejnych okółków w kwiecie typu dzikiego wyglądu więc następująco:
A (kielich) A + B (korona) B + C (pręciki) C (owocolistki)
Wyszukiwarka
Podobne podstrony:
17086 skanowanie0014 (22) Rys.15. Zależność stopnia pochłaniania światła A, stopnia zaciemnienia spa
skanowanie0018 (64) Rys. 15-2. Po*e rnof ogtoa»yC2ne w SMfl ttar«ionyrr metodą Feiigana Jądra Koi&nb
CCI20101218�021 22 Rys. 15. Schemat przyrządu AEG służącego do hydrostatycznego ciągnienia drutu: 1
skanowanie0019 (60) Rys. 15-3. Dwa nakrzyżlacjla ulistnione pędy jałowca, w Których widoczna jest zm
skanowanie0042 (24) Rys. 15-11. (A) Liść podwójnie z ozony GfeoCscfóa z efektami homeotycznymi. pole
badwłasn0011 22 I Rys. 15* Pod wpływem przyłożonego obciążenia próbka odkształca się, a równocześnie
więcej podobnych podstron